While Google and Tesla are working on self-driving cars, Terrafugia’s TF-X has its sights set much higher, literally.
вторник, 29 сентября 2015 г.
6 увлекательных YouTube-блогов обо всем на свете
Иногда у нас абсолютно не хватает времени, чтобы записаться на курсы или даже пройти полноценный онлайн-курс. Но мы все равно стремимся развиваться и узнавать что-то новое. Отличный помощник в этом – YouTube. Ведь здесь полно интересных и познавательных видео о физике, географии, истории.
Они намного проще и легче воспринимаются, чем онлайн-курсы, и станут отличным способом отвлечься даже для тех, у кого был непростой рабочий день. Кроме того, это отличный способ потренировать английский язык.
1. Mental Floss
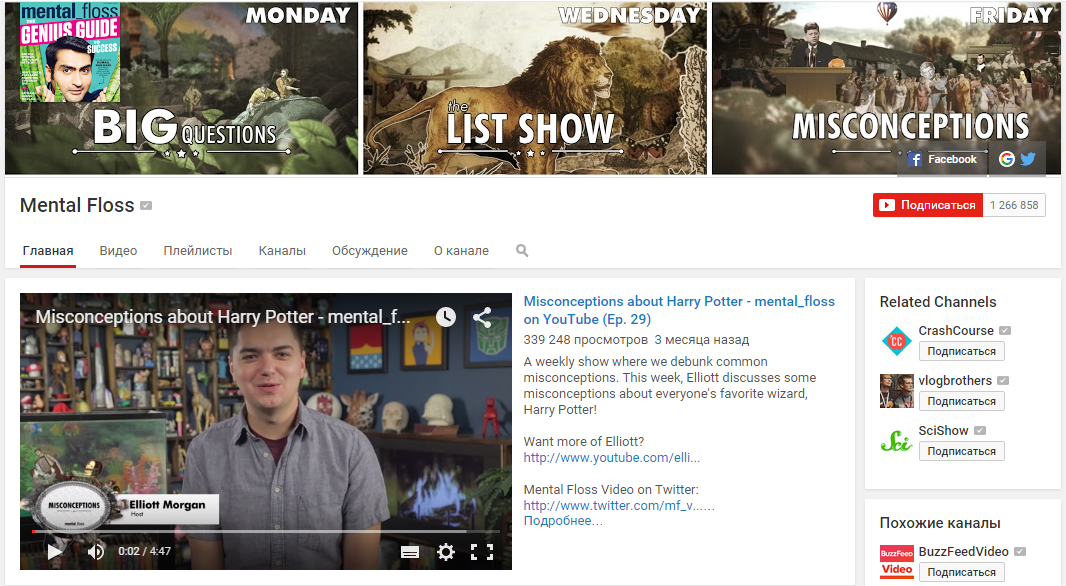
Большой развлекательный канал о разнообразных интересных фактах и явлениях из разных сфер. Ролики сделаны в формате коротких видео с набором фактов, посвященных определенной теме. На канале представлено сотни мини-лекций о самых разнообразных вопросах – от видео о происхождении китайских имен до вопроса, почему люди наклоняют голову, когда пытаются лучше понять собеседника.
Условно канал разделен на три направления – интересные факты (например, 27 странных мировых рекордов, 31 факт о свиньях или 24 непонятных научных исследования), ошибочные концепции (например, представления об истории, социальных медиа или космосе) и т. н. большие вопросы (к примеру, почему мы стучим зубами, когда мерзнем или почему клей не прилипает к банке, в которой находится). Канал обновляется еженедельно и собрал уже больше миллиона подписчиков.
2. CGPGrey
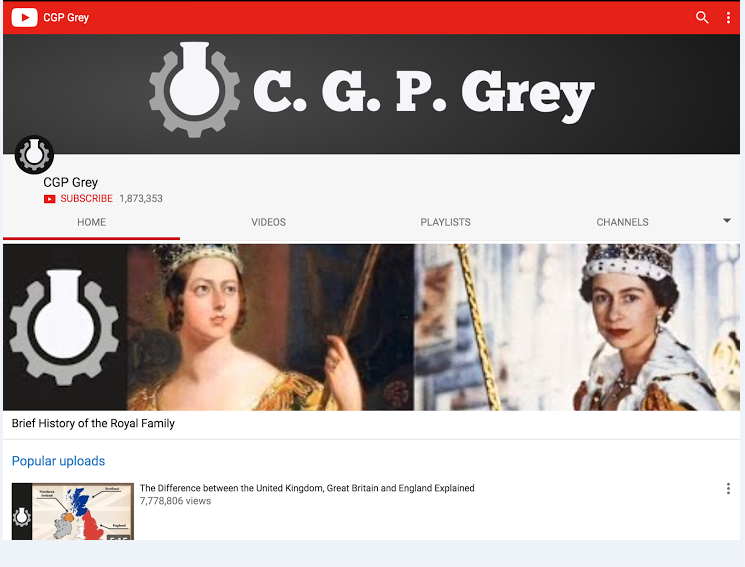
Еще один канал о разнообразных фактах из истории, литературы, географии и политологии. Каждое видео посвящено одному вопросу. Например, как появился Ватикан, как проходят выборы в Великобритании, во сколько бюджету обходится содержание британской королевской семьи или кто есть кто в фэнтезийном мире Толкиена.
Благодаря легкой ироничной подаче и структурированной информации, получается быстро разобраться в сложных и запутанных темах.
3. Periodic Videos
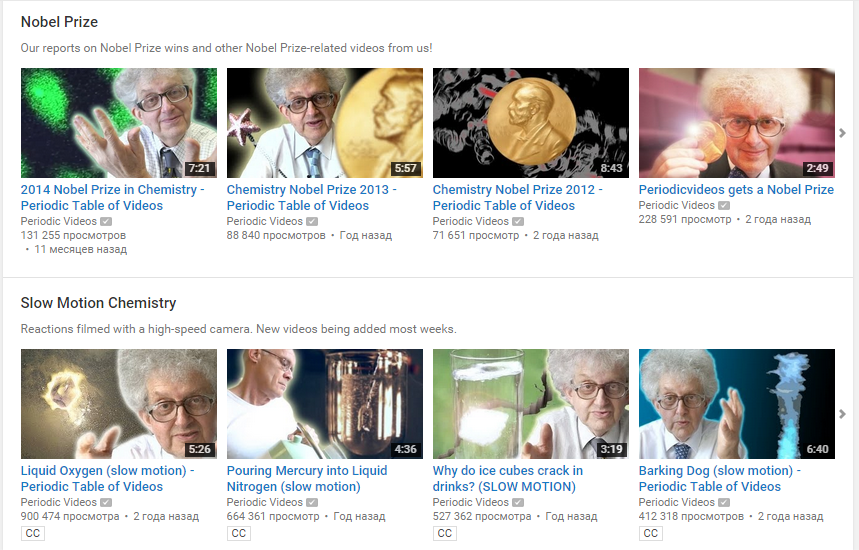
На YouTube можно найти много курсов о химии, однако фишка этого курса в том, что он начинается с самых основ – со знакомства с каждым химическим элементом. Каждому из известных 118 элементов посвящено отдельное видео. В очень простой форме рассказывается о свойствах элемента. Кроме того, каждую неделю на канал добавляются видео о научных экспериментах, открытиях и новостях.
4. Sonia’s Travels
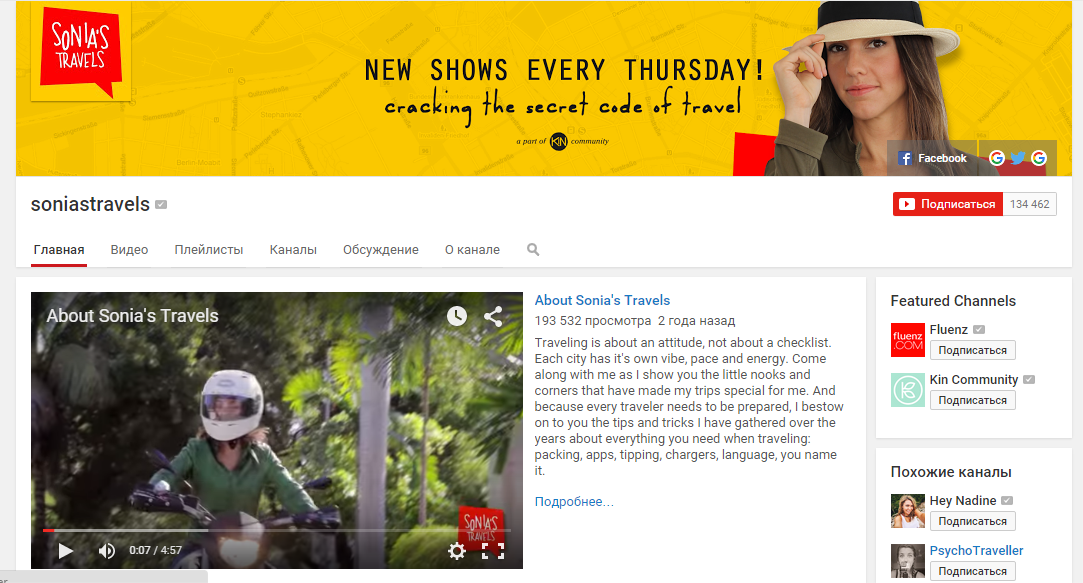
Очень увлекательный и вдохновляющий видеоблог о путешествиях. Автор блога Соня Джил путешествует по разным городам мира и рассказывает о популярных достопримечательностях, малоизвестных живописных местах, а также о жизни людей и своих впечатлениях. Кроме историй о новых странах, на блоге можно найти полезные советы на разные темы, связанные с путешествиями, например, как путешествовать одному, как облегчить багаж или привести в форму свою одежду без утюга, как комфортно поселиться в хостеле и как заговорить с незнакомцем. Обновления на канале появляются каждую неделю. Безусловный плюс видеоблога – качественно снятая и смонтированная картинка.
5. Minute Physics
Это один из самых популярны видеоканалов о физике на YouTube, собравший уже почти 2 миллиона подписчиков. Создавая видео, авторы этого канала отталкиваются от известного выражения: «Если ты не можешь объяснить просто, значит, ты недостаточно хорошо разобрался в этом». Здесь вы найдете сотни видео с объяснениями разных физических понятий и явлений. Каждое видео посвящено одной теме. Создатели канала фокусируют внимание, как на теории, например, рассказывая что такое гравитация или темная материя, так и на разъяснении бытовых ситуаций, с которыми мы сталкиваемся в повседневной жизни с точки зрения физики. Например, можно узнать, что происходит, когда вы едете на велосипеде или что лучше делать под дождем – идти или бежать с точки зрения физики.
6. Speak English with Misterduncan
«Английский без единого вызова к доске!», – такой девиз этого канала. Видео созданы таким образом, что, скорее, похожи на разговор о языке, чем сухое изучение языка. Автор канала записывает монологи в разнообразных языковых ситуациях, отвечает на вопросы о языке, подробно разъясняя разные выражения. Уроки сопровождаются комедийным и местами абсурдным видеорядом. Канал будет интересным и полезным для тех, кто интересуется английским и хотел бы углубить свои знания, а также лучше почувствовать специфику английского языка. Создатель курса утверждает, что видео подойдут абсолютно для любого зрителя, но поскольку все видео записаны на английском, то базовое знание языка все же необходимо.
5 Craziest Bridges of The World
#1 Japan's Eshima Ohashi bridge: The kind of bridge you'd expect in Grand Theft Auto
At 44 meters high, the bridge promises a sharp plummet down, and you can imagine the Japanese silently cursing "Not this crap again" as they reach the top of the bridge and double check their seat belts.

#2 Root bridge in Cherrapunji: This bridge is alive!
Since Cherrapunji is one of the wettest places in India, ordinary wood bridges would collapse here. To guide the roots holding the bridge together, they use hollowed out betel nut trunks. So rocks, roots, and mud make 100 feet bridges that can support 50 people (or over 500 kilos). Oh, and some of these bridges are already over 500 years old!

#3 Henderson Waves - Singapore comes here to get legally high
The highest you can get on a bridge in Singapore is here, at Henderson Waves bridge, 36 metres above ground. Which makes sense - it looks like stoners built it. For some odd reason, the one thing Singaporeans love to do here is take jumping selfies.

#4 Don't look at the Nanpu Bridge in Shanghai for too long. You'll get dizzy.
Those loops make you say WOW. And kind of afraid if your car breaks down and you stall the traffic here - last clocked at 120,000 vehicles per day in 2006. The Chinese have described it as a dragon laying across the river. Which is expected - these guys love their dragons.

#5 The Rolling Bridge: London is so Brit
While most of the bridges on this list are crazy, London's Rolling Bridge is just practical and oh-so-Brit. It tidily rolls up when you don’t need it, and can get you across the Grand Union Canal when you do.
And then it curls back again so boat traffic can pass by. Hydraulic pistons power the steel and timber footbridge's movement.
And then it curls back again so boat traffic can pass by. Hydraulic pistons power the steel and timber footbridge's movement.

понедельник, 28 сентября 2015 г.
Почему перелетные птицы возвращаются весной обратно?

На этот вопрос можно корректно ответить по крайней мере с трех разных позиций. Эти ответы будут дополнять друг друга и потому в равной степени важны. Во-первых, каков механизм этого явления? Во-вторых, зачем птицы это делают — в чем значение (функция) этого поведения? И, наконец, как вообще произошло так, что птицы куда-то улетают, а потом возвращаются (то есть каково происхождение и эволюция этого явления)? Ниже вкратце рассмотрим эти три аспекта.
Как?
Если содержать перелетных птиц в неволе, то в период обычных сезонных миграций они испытывают беспокойство. Такое состояние было названо миграционным. В это время, например, можно наблюдать нетипичную активность по ночам. А связано это с тем, что мелкие птицы летят в основном ночью. То есть они как бы пытаются реализовать свою тягу мигрировать даже в том случае, если им (в неволе) этого сделать не дают.
Более того, птицы стараются ориентироваться в ту сторону, куда они в норме должны лететь. Эта особенность широко используется при изучении ориентации птиц с помощью так называемых круглых клеток, или клеток Крамера, названных в честь немецкого орнитолога Густава Крамера (Gustav Kramer). В таких клетках (круглой формы) по периметру расположены жердочки, а одна жердочка находится в центре клетки. При прыжках птице удобнее прыгать с центральной жердочки на одну из периферийных. По тому, куда ориентирована (по сторонам света) наиболее часто посещаемая периферийная жердочка определяют, в какую сторону птица «хочет» мигрировать.
Итак, желание мигрировать на юг (осенью) или возвращаться домой (весной) проявляется у птиц даже тогда, когда им не дают это сделать. То есть миграционное состояние, по сути дела, явление инстинктивное. Запускается оно у наших птиц главным образом соотношением между темным и светлым временем суток (так называемым фотопериодом). Определенное значение этого параметра является своеобразным триггером миграций. Это показано, в том числе, экспериментально.
Зачем?
Теперь посмотрим, зачем птицам надо возвращаться домой. В чем смысл (функция) этого? Как это помогает им выживать? Ведь чтобы сформировался инстинкт, о котором шла речь в предыдущем подразделе, он должен иметь какую-то ценность — иначе он бы просто не возник.
В жизни птиц можно выделить несколько периодов. Они повторяются каждый год, поэтому обычно говорят о годовом цикле. В типичном случае годовой цикл выглядит так: гнездование, линька, осенняя миграция, зимовка, весенняя миграция, снова гнездование и далее «по списку». Все названные периоды важны, но особое значение имеет гнездовой. В это время птицы выводят потомство, от них требуется масса дополнительных затрат — как времени, так и энергии. Поэтому успешно размножаются лишь те особи, которые делают это в благоприятных для них местах, к которым они лучше всего приспособлены.
Почему же наши птицы обычно не гнездятся, например, в тропиках? Здесь две главных причины. Во-первых, они недостаточно хорошо приспособлены к тамошним условиям. То есть они могут жить там, добывать себе пищу, даже петь, но на большее их не хватает. Трудно найти подходящее место для гнезда, трудно выкормить птенцов и т. п. А во-вторых, в тропиках масса местных оседлых видов, которые «переигрывают» мигрантов в конкурентной борьбе — как в прямой (например, за гнездовые убежища), так и в косвенной (за пищу).
Но бывает и так, что наши северные птицы и где-то далеко на юге находят подходящие для себя условиях и остаются там гнездиться. В некоторых случаях это может даже со временем привести к появлению новых форм. Хороший пример — утка-кряква (Anas platyrchynchus, рис. 1), обычная в средней полосе России, и в том числе в черте Москвы. А кроме этого, она гнездится по всей Северной Америке и Евразии, от тундр до субтропиков. Так что этот вид очень пластичен. Поэтому, быть может, и не удивительно, что некоторые популяции, попав в ходе миграций на тропические острова, остались там жить и стали оседлыми.

Рис. 1. Обыкновенная кряква (слева самка, справа самец). Фото с сайта ru.wikipedia.org
|
Сейчас такие формы считают даже отдельными (но близкими) видами. Это гавайская кряква Anas (platyrhynchus)wyvilliana и лайсанский чирок Anas (platyrhynchus) laysanensis, оба вида — с Гавайских островов (рис. 2).

Рис. 2. Гавайская кряква (слева) и лайсанский чирок. Самцы и самки у этих видов не различаются и напоминают самку обыкновенной кряквы. Фото с сайта birdlife.org
|
Бывают и еще более интересные исключения. Одно из них — черный шелковистый свиристель (Phainopepla nitens, рис. 3), обитающий в Северной Америке. Эта птица умудряется гнездиться дважды в году. Весной она выводит птенцов в штате Калифорния. А к осени мигрирует в штат Колорадо. Здесь она гнездится еще раз. Такое гнездование в двух разных местах — уникальный случай у птиц. Так что, как это вообще характерно для зоологии, имеются лишь общие тенденции или правила с множеством разных исключений.

Рис 3. Черный шелковистый свиристель (Phainopepla nitens). Слева самец, справа самка. Фото с сайта en.wikipedia.org
|
Наконец, надо коротко рассказать, зачем птицы вообще улетают зимой в теплые края. Главнейшая причина — нехватка пищи. Поэтому в первую очередь улетают те виды птиц, которые питаются открыто живущими насекомыми. Зимой такую пищу, конечно же, не найти. Так что они мигрируют, можно сказать, вынужденно. Те же виды, которые могут найти себе корм и зимой, остаются в наших краях. Это, например, синицы, ловко выискивающие спящих насекомых в различных щелях и разнообразящие свой рацион семенами. Или большой пестрый дятел (Dendrocopos major), питающийся зимой семенами ели и сосны.
Почему?
Но почему птицы, которые гнездятся в северных широтах, а зимуют в тропических, делают именно так, а не иначе. Почему бы им, например, не гнездиться зимой в тропиках, а летом не отправляться отдыхать на север? Для ответа на это необходимо рассмотреть также эволюционный аспект. А именно — историю расселения видов.
Дело в том, что многие виды наших птиц имеют южное происхождение. Все они — выходцы из Африки или Южной Азии. В ходе своей эволюционной истории они постепенно расселялись из этих областей. Формировались новые популяции и виды, адаптирующиеся к новым, более северным, условиям. Столкнувшись в новых условиях с неблагоприятной обстановкой зимой, эти птицы вынуждены были мигрировать на юг. И путь этот пролегал в те районы, откуда эти виды изначально произошли. Своего рода историческая память. Поэтому существует известная аналогия, что путь миграции в общих чертах повторяет путь расселения вида. Конечно, не стоит точно связывать район зимовок и район, откуда началось расселение. Соответствие здесь есть, но оно примерное. Так, если вид зимует в тропической Азии, можно говорить о его азиатском происхождении, но вовсе не обязательно тропическом.
Районы зимовок могут оставаться консервативными, даже если это и не очень удобно. Такова, например, ситуация с овсянкой-дубровником (Emberiza aureola) — азиатским видом, недавно расселившимся в Европу, вплоть до Прибалтики. Конечно, европейским птицам короче было бы летать на зимовку в Африку, тем не менее они «по старинке» летают в юго-восточную Азию — как и птицы из Сибири и Дальнего Востока (рис. 4).

Рис. 4. Гнездовой (красный) и зимовочный (зеленый) ареалы овсянки-дубровника. Схема составлена по материалам сайта xeno-canto.org, фото А. С. Опаева
|
Дубровник лишь недавно стал гнездиться в Европе. Но большинство других видов, более давних выходцев из Азии, со временем поменяли места зимовок. Европейские популяции стали проводить зиму в Африке — что, очевидно, и ближе, и удобнее.
Таким образом, история вида также важна для понимания того, как он ведет себя сейчас. Но ни один из трех аспектов (механизм, функция, эволюция), взятый в отдельности, не может ответить на поставленный вопрос. И лишь в совокупности они рисуют цельную картину того, зачем и почему птицы возвращаются весной назад.
Ответил: Алексей Опаев
Почему черепахи долго живут?

Если ответить совсем коротко, то черепахи живут долго, потому что они а) холоднокровные и б) большие. Поясним эти два ответа.
Жизнь поддерживается химическими реакциями. Чтобы существо оставалось живым, химические вещества в нем постоянно должны превращаться друг в друга, или, как еще говорят, обмениваться. Скорость этих превращений, или обмена веществ, напрямую связана с тем, насколько активным может быть животное, сколько пищи ему нужно употреблять, сколько ему нужно спать и сколько оно проживет.
Скорость обмена веществ зависит от размера животного. У маленьких животных поверхность тела большая относительно объема, а с увеличением размера животного площадь поверхности растет медленнее, чем его объем. Например, у слона на 1 кубический сантиметр объема тела приходится примерно 0,03 квадратных сантиметров поверхности тела, а у хомячка — целых 0,6, то есть в 20 раз больше. Через эту большую поверхность улетучивается тепло, которое хомячок старательно вырабатывает, поэтому хомячку приходится есть и сжигать намного больше питательных веществ в расчете на единицу массы тела, чем слону. Обмен веществ хомячка с неизбежностью идет быстрее, потому что он много ест и вырабатывает много тепла, а от переработанной пищи быстро избавляется и немедленно поглощает новую.
Животные с быстрым обменом веществ более активные (когда не спят), потому что у них вырабатывается много энергии и потому что им всё время нужна новая пища. При этом им нужно больше спать, из-за того что в их мозге быстрее накапливаются токсичные побочные продукты работы нейронов, для избавления от которых, по-видимому, необходим сон. Если мелкие грызуны спят до 20 часов в сутки, то слоны — всего 3–5 часов.
Позвоночные животные с медленным обменом веществ (то есть крупные животные) живут дольше животных с быстрым обменом веществ — это известная закономерность. Рекордсмены по продолжительности жизни среди позвоночных — это киты, слоны и галапагосские черепахи, тоже довольно крупные и растущие, по-видимому, в течение всей жизни.
Почему с медленным обменом веществ можно прожить дольше, чем с быстрым, это не такой простой вопрос. Ответ как-то связан с накоплением повреждений и старением, которые происходят быстрее у животных с быстрым обменом веществ, но не совсем напрямую. К примеру, мелкие грызуны голые землекопы знамениты своим отсутствием признаков старения и устойчивостью к раку (который может возникать из-за самых разных повреждений). Но даже без старения и с замечательной устойчивостью к повреждениям эти животные живут не дольше 30 лет. Это намного больше, чем средняя продолжительность жизни у других животных такого размера (которая составляет около двух лет), но все-таки существенно меньше средней продолжительностей крупных животных, которые при этом не свободны от старения. В теории, голые землекопы могли бы жить до тех пор, пока их кто-нибудь не съест или с ними не произойдет какой-то другой несчастный случай, но все-таки иногда они умирают, не дожив до этого.
Разумнее всего выглядит предположение о балансе рождений и смертей, который должен поддерживаться в популяции. В какой-то момент любая особь должна погибнуть, чтобы освободить жизненное пространство для нового, более приспособленного поколения. Но она должна делать это в определенный момент, сообразный с ее скоростью размножения. У животных с быстрым обменом по сравнению с остальными жизнь проходит как будто в ускоренной съемке — они проживают ее быстрее, но и более активно, оставляя большое количество потомков за достаточно краткий срок. Поэтому они быстро справляются со своей задачей участников популяции и быстро же освобождают место для нового поколения. Слоны или другие крупные животные размножаются намного реже мелких животных и приносят меньше потомства. Их жизнь как бы замедлена, так что им необходимо жить дольше, чтобы всё успеть.
Точно так же замедлена жизнь у холоднокровных животных по сравнению с теплокровными. Холоднокровные животные не тратят ресурсов на поддержание постоянно высокой температуры и тела, а значит, и высокой скорости обмена. Поэтому скорость обмена у них такая, как повезет: если ящерица погреется на солнышке, она будет активнее, а если нет, то ей придется немного замедлиться. Но в среднем скорость обмена веществ у холоднокровных животных всегда ниже, чем у теплокровных, потому что и солнечно бывает не всегда, и нагреться на солнце до таких температур тела, как у млекопитающих или птиц, обычно нелегко. Поэтому жизнь холоднокровных животных еще более замедлена, чем у теплокровных.
Черепаха — холоднокровное животное, и черепахи действительно живут довольно долго. А вот большие черепахи — жители Галапагосских островов, совмещают замедленность обмена крупных животных и холоднокровных животных. Такое удачное сочетание позволило им поставить рекорд среди позвоночных животных (если брать хорошо задокументированные свидетельства) — 177 лет жизни.
Но черепахам не стоит завидовать, потому что, как уже говорилось, их скорость метаболизма отличается от нашей. Скорость метаболизма связана с ощущением субъективного времени: чем она выше, тем медленнее в восприятии животного меняется мир вокруг. Эту скорость индивидуального времени можно узнать, измерив минимальную частоту вспышек света, начиная с которой свет начинает восприниматься как непрерывный, — так называемую критическую частоту слияния мельканий (КЧСМ).
|

У человека это число в среднем равно 60 вспышкам в секунду, а у черепахи — 15. Это означает, что по ощущениям черепахи время идет в 4 раза быстрее, чем для человека: за время, за которое человек успевает заметить 4 мелькания, черепаха может заметить только одно.
Легче это представить на примере с мухами, у которых КЧСМ в 4 раза больше, чем у людей, 240 мельканий в секунду. Человеку кажется, что он быстро замахивается на муху газетой, но для мухи его движение выглядит очень медленным, поэтому по ней почти невозможно попасть. А снаружи все выглядит наоборот — животное, которому кажется, что время быстро летит, движется медленно.
То есть 177 лет для черепахи — это не так уж много.
Так что первое, что можно извлечь из этой истории, — что черепахам не стоит завидовать. Второе, немного менее очевидное, — что уменьшение калорийности пищи может увеличить продолжительность жизни. Действительно, чем больше питательных веществ мы перерабатываем, тем выше метаболическая нагрузка на наш организм. Она не проходит бесследно, особенно у тех животных, которые, в отличие от голых землекопов, и стареют, и не обладают невероятной устойчивостью к повреждениям. Особенно обидно перерабатывать лишние калории, которые большинству современных людей, ведущих сидячий образ жизни, не нужны. Эксперименты по ограничению калорийности пищи проводились на мышах и крысах, а также некоторых рыбах и обезьянах, и дали неплохие результаты. У мышей, например, среднюю и максимальную продолжительность жизни удавалось увеличивать на 30–50%, уменьшая калорийность диеты на 55–65% (C. M. McCay, Mary F. Crowell, and L. A. Maynard, 1935. The Effect of Retarded Growth Upon the Length of Life Span and Upon the Ultimate Body Size: One Figure).
Ответила: Юлия Кондратенко
Человек из Диналеди — новый вид примитивных людей

Реконструкция облика человека из Диналеди; возможно, лицо будет несколько «подправлено» в дальнейшем, когда найдутся более полные черепа (пока черепов с лицевой частью не откопали), а цвет глаз и волос изменят, если (или когда) научатся читать ДНК из столь древних остатков. Реконструкция National Geographic Society. Изображение с сайта nbcnews.com
|
Большая международная команда антропологов под руководством Ли Бергера описала новый вид гоминид, которого она отнесла к роду Homo. Открытое местонахождение является уникальным по количеству костей (поднято на поверхность полторы тысячи костных единиц, из них только десяток не принадлежит гоминидам) и полноте скелетов. В анатомии представителей нового видаH. naledi сочетаются признаки людей и австралопитеков, однако человеческих черт всё же больше. Для понимания эволюционной истории важно, что небольшому мозгу приданы маленькие зубы и развитые, вполне человеческие, руки и стопы. Это означает, что «человеческая» эволюционная линия, характеризующаяся такими признаками, как двуногость, трудовая деятельность, пониженная агрессивность и развитые социальные отношения, не обязательно подразумевает укрупнение мозга. К сожалению, пока не удалось датировать остатки, а также определить, как все эти люди погибли и попали в труднодоступную камеру пещеры.
В послужном списке южноафриканского антрополога Ли Бергера (Lee Berger) уже имеются несколько достойнейших открытий. Напомню их. В 2006 году он переисследовал знаменитый череп ребенка из Таунга и доказал, что ребенок стал добычей крупной хищной птицы (см.: Юные австралопитеки были добычей хищных птиц, «Элементы», 28.09.2006). Через два года Бергер публикует работу с описанием захоронения низкорослых людей на острове Палау, рассуждая попутно о карликовости островных популяций людей, в том числе, и с острова Флорес (На островах в Тихом океане обнаружены кости карликов», «Элементы», 17.03.2008). И затем в 2010 году последовало открытие нового вида австралопитеков, получивших наименование Australopithecus sediba (Australopithecus sediba — австралопитек, похожий на человека, «Элементы», 15.04.2010).
Из двух новых публикаций в журнале eLIFE читатель узнает, чем занимался этот неутомимый и азартный антрополог в течение прошедших четырех с половиной лет. Как выяснилось, он вел раскопки в одной из пещер в ЮАР в 50 км от Йоханнесбурга. В течение двух полевых сезонов — осенью 2013-го и весной 2014-го — его команда исследовала человеческие останки, найденные двумя спелеологами в одной из камер пещеры. Пещера имеет очень сложную форму с несколькими залами и узкими вертикальными и горизонтальными тоннелями-«шкуродерами», пройти которые полностью удалось, скорее всего, впервые.
Чтобы протиснуться в узкие лазы, имеющие в некоторых местах ширину 20–25 см, пришлось поискать для работы худых геологов (понятно, что для этого лучше всего подошли стройные юные девушки). Так или иначе, но пещерные тоннели были пройдены, и в последней камере, именуемой Диналеди, была найдена богатейшая коллекция человеческих останков. Наверх для исследования было поднято около 1500 костей и зубов, все достойной сохранности, некоторые скелеты удалось собрать практически полностью. Среди них все принадлежат гоминидам, и только 6 птичьих костей и несколько плохо определимых остатков грызунов.
По числу костных остатков, их концентрации и сохранности это местонахождение уникально. За прошедшие века захоронение не потревожили ни звери, ни птицы, ни туристы, и оно сохранилось в своем первоначальном состоянии, покрываясь постепенно тонкими наносами глинистой пыли. Так что в прошедшие четыре года для Ли Бергера нашлось весьма интересное занятие. Нужно было понять, чьи эти кости, как они туда попали и из какого они времени.
Ответ на первый вопрос такой: все кости бесспорно принадлежат гоминидам одного вида, и это новый вид людей — Homo naledi. Вид назван по имени пещеры Диналеди (Dinaledi, см. Rising Star Cave), а вот отнесение к роду Homo потребовало анализа морфологических признаков скелета и сравнения со всеми имеющимися гоминидами и человекообразными обезьянами. Благо материала для сравнения было предостаточно, и он тщательно описан (профессиональным антропологам будет интересно посмотреть на сравнительную сводкуморфологических черт Homo naledi и других гоминид, непрофессионалам непросто будет разобраться в латинских наименованиях костных элементов).
Люди из Диналеди оказались небольшого роста (144–148 см) и веса (около 39–45 кг), с объемом мозга 465–550 см3, некрупными зубами, в том числе и маленькими клыками, жевательные поверхности коренных зубов у них относительно простые. Ходили эти люди на двух ногах, уже вполне человеческих, если не считать небольшого плоскостопия. На таких ногах, более длинных, чем у австралопитеков, можно было совершать дальние походы. Кисти рук у них тоже были больше похожи на человеческие — с удлиненным большим пальцем и подвижным запястьем, правда пальцы кривоваты. Удивительно при этом, что их ребра и плечи соответствуют австралопитечной морфологии, то есть годятся для лазанья; кстати, и искривленные фаланги пальцев тоже напоминали пальцы австралопитеков. Также и крепления бедер и вся тазовая часть скорее напоминают австралопитеков. Так что перед нами вид гоминид, сочетающих черты людей и австралопитеков.

При небольшом размере мозга, сопоставимом с размером мозга австралопитеков и карликовых людей с острова Флорес, у Homo naledi оказались совсем небольшие зубы, какие характерны для людей современного типа. Раньше предполагалось, что тенденция к увеличению мозга связана, в том числе, с уменьшением размера зубов. Это считалось симптомом снижения агрессии в группах и становления новой социальной системы.H. naledi не слишком хорошо укладывается в эту схему. Рис. из обсуждаемой статьи L. Berger et al., 2015
|
Обсуждая мозаику признаков, антропологи обращают внимание на весьма важную деталь. Эти люди имели руки, способные к мелким и точным манипуляциям, стопы, приспособленные для двуногого хождения, а также небольшие зубы, свидетельствующие о снижении агрессии. Все эти качества вроде бы предполагают наличие относительно крупного мозга. У этих же людей, напротив, мозг оставался маленьким. Означает ли это, что весь «человеческий комплекс» — двуногость, трудовая деятельность и социальность — не требует большого мозга? Это исключительно важный вопрос для понимания эволюции человека, на который еще предстоит ответить.
Другой вопрос, так и не нашедший ответа, — датировки захоронения. В этом местонахождении кости залегают в двух слоях, которые были отчасти перемешаны из-за периодического обводнения камеры пещеры. Поэтому датирование по изотопам урана дает слишком большую ошибку, поскольку материал из слоев перемешан. Датировки по изотопам углерода невозможны, так как на временных интервалах выше первых сотен тысяч лет они не срабатывают. Косточки грызунов, по которым в принципе можно было бы прикинуть возраст костеносных слоев, слишком повреждены и плохо сохранились и потому не определяются как следует. Ученые надеются, что в будущем им удастся подобрать адекватную методику для датирования людей из Диналеди. А пока что их место на генеалогическом древе гоминид — где-то между австралопитеками и людьми. Датировки помогут определить его более точно. Тогда можно будет понять, как расположить новобранцев из Диналеди по отношению к хабилисам, эректусам и H. rudolfensis.
Еще одна загадка этого местонахождения — как попали все эти люди в глубокую и трудно проходимую пещеру. Геологи утверждают, что других входов в пещеру и следов обвалов нет. Это означает, что в древности, как и сегодня, во внутренние камеры пещеры попасть было нелегко. Тем не менее все признаки — положение скелетов, повреждения костей, нахождение почти полных скелетов — говорят о том, что останки (или еще живые люди) доставлены были в пещеру целиком, а не по частям. Так что версию с хищниками, таскавшими в укромное место свою трапезу, можно отбросить. Также пришлось оставить и гипотезу о лавинах и потоках, ставших причиной смерти и смывших погибших в глубины пещеры. В этом случае на костях остались бы характерные следы.
Вряд ли пещеры были местом ритуальных захоронений: тогда можно было бы ожидать большую долю захоронений пожилых людей. Но среди умерших пожилым можно считать только одного человека; другие погибшие (по чьим костям определялся возраст) были моложе: трое детей, четверо подростков и четверо взрослых. Хотя ученые и не исключают гипотезу о намеренном захоронении и даже всячески ее пропагандируют. Ведь детская смертность, напоминают они, во все времена была исключительно высока. Эта гипотеза, подтвердись она тем или иным способом, даст начало интересным выводам о древности ритуальных традиций. Пока что, однако, говорить об этом рано.
Возможно также, что причиной гибели стал неожиданный обвал или другая катастрофа, и в результате целая группа людей оказалась погребена в смертельной ловушке. Опять же, этот вопрос остается открытым.

Схема пещеры Диналеди, где откартировано положение видимых остатков костей и площадь раскопа. Как следует из схемы, раскопки только начались. Рис. из обсуждаемой статьи P. Dirks et al., 2015
|
Площадь раскопок в пещере далеко не покрывает всего костеносного слоя. Вероятно, в ближайшем будущем команда антропологов, откопав еще сотню-другую костей, ответит на некоторые из вопросов. Но скорее всего, как это обычно бывает, с ответами появятся другие вопросы. Тем интереснее нам ждать продолжения истории.
Мутирующее человечество: что мы узнали о своих мутациях за 15 лет геномной эры

Рис. 1. Скорость мутагенеза у разных организмов. По вертикальной оси — темп нуклеотидных замен (на миллиард пар оснований за поколение), по горизонтальной — размер генома (в миллионах пар оснований). Левый график — вирусы и прокариоты,правый — эукариоты (на правом графике также показаны усредненные значения для бактерий и архей). Красная линия соответствует уровню мутагенеза, равному одной мутации на геном за поколение. Видно, что у вирусов и прокариот чем больше геном, тем ниже темп мутагенеза, тогда как у эукариот наблюдается обратная зависимость. Видно также, что млекопитающие характеризуются экстремально высоким числом мутаций на геном за поколение. Eubacteria — бактерии, Archaea — археи, Double-stranded DNA viruses, Single-stranded DNA viruses — вирусы, содержащие дву- и однонитевую ДНК, RNA viruses — РНК-содержащие вирусы, Mammals — млекопитающие, Invertebrates — беспозвоночные, Plant — растение, Unicellular eukaryotes — одноклеточные эукариоты. Изображение из статьи: Lynch M., 2010. Evolution of the mutation rate // Trends in Genetics. V. 26. P. 345–352 (с изменениями)
|
За 15 лет, прошедших со времени первого чернового прочтения человеческого генома, наши знания о мутациях, то и дело возникающих в наших генах, многократно увеличились, а методы их изучения стали намного эффективнее и разнообразнее. В обзорной статье американских генетиков, опубликованной в журнале Science, подводятся итоги изучения мутационного процесса у Homo sapiens за первые полтора десятилетия «геномной эры».
Мутагенез — одно из самых фундаментальных биологических явлений. Все без исключения живые существа подвержены мутациям. Случайные изменения в последовательности нуклеотидов ДНК возникают как во время репликации (размножения) молекул ДНК, так и в промежутках между актами репликации (например, под действием ультрафиолетового излучения и других мутагенов). Большинство мутаций сразу исправляется специальными системами починки (репарации) ДНК, но даже самые сложные и совершенные системы репарации не обеспечивают стопроцентную защиту.
Темп мутагенеза сильно различается у разных организмов, однако ни у одного существа — от вирусов до человека — он не равен нулю. Этот факт может показаться тривиальным, но на самом деле он требует объяснений, и вот почему.
Как правило, среди ненейтральных (влияющих на приспособленность) мутаций гораздо больше вредных, чем полезных. Так получается по чисто вероятностным причинам (см. Принцип Анны Карениной). Следовательно, чем ниже темп мутагенеза, тем выше, при прочих равных, будет средняя приспособленность потомков данного организма. Поэтому отбор, казалось бы, всегда должен способствовать снижению темпов мутагенеза: под его действием должны развиваться всё более точные системы репликации и репарации. Почему же эти системы так и не стали абсолютно безошибочными?
Причин, скорее всего, две. Первую можно условно назвать «экономической». Сверхточные системы репликации и репарации, скорее всего, были бы слишком дороги: громоздки, энергоёмки, их работа замедляла бы репликацию или имела какие-то другие вредные побочные эффекты. К тому же чем точнее работают эти системы, тем слабее давление отбора, направленного на их дальнейшее усовершенствование.
Вторая причина состоит, конечно же, в том, что полезные мутации тоже нет-нет, да и происходят (см.: Ранние этапы адаптации предсказуемы, поздние — случайны, «Элементы», 03.03.2015). Поэтому, хотя средняяприспособленность потомков мутирующих организмов всегда будет ниже, чем у немутирующих, разброс приспособленности и максимальная приспособленность у первых будут выше. Поэтому во многих ситуациях, особенно при изменениях среды, явное преимущество будет на стороне мутирующих организмов не только в долгосрочной перспективе (тысячи и миллионы лет), но и в краткосрочной (в масштабе одного или нескольких поколений). Для некоторых организмов прямой вред слишком медленного мутирования удалось показать экспериментально (см.: Вирусы-мутанты помогают друг другу в борьбе за выживание, «Элементы», 14.12.2005).
Так или иначе, мутации неизбежны, вредных среди них гораздо больше, чем полезных, и всему живому приходится с этим мириться (и, более того, к этому приспосабливаться). Мы, люди, не только не являемся исключением, но даже, вместе с остальными млекопитающими, сильно опережаем большинство живых существ по темпам мутагенеза в расчете на особь за поколение (рис. 1).
Геном Homo sapiens впервые был вчерне прочтен около 15 лет назад (см. Проект «Геном человека»). С этого момента началась новая эра в изучении мутационного процесса у нашего вида. Благодаря стремительному развитию технологий секвенирования и анализа геномных последовательностей сегодня мы знаем неизмеримо больше, чем в начале XXI века, о темпе мутаций, о закономерностях их распределения по геному, об их роли в различных патологиях (включая рак) и о других особенностях нашего мутагенеза, имеющих как теоретическое, так и практическое значение.
В свежем выпуске журнала Science опубликована подборка статей, посвященных этой животрепещущей теме. Большинство из них касаются узкоспециальных, в том числе медицинских вопросов, но в одной, написанной генетиками из Вашингтонского университета в Сиэтле, дан общий обзор того, что мы узнали о своих мутациях за последние 10–15 лет.
1. Темп мутаций «зародышевой линии». Все мутации можно подразделить на соматические (происходящие в соматических, то есть не половых клетках организма на разных стадиях развития) и мутации «зародышевой линии» (germline mutations), меняющие геном половых клеток и передающиеся по наследству потомкам. С медицинской точки зрения крайне важны оба типа мутаций, с эволюционной — вторые, конечно, важнее.
Первые оценки скорости возникновения мутаций в зародышевой линии у человека были сделаны задолго до геномной эры, однако их точность была невелика. Сегодня для этого используют несколько подходов. Один из них — изучение родословных с целью подсчета вновь возникающих мутаций с четким фенотипическим эффектом и высокой пенетрантностью (то есть таких мутаций, которые меняют фенотип, во-первых, строго определенным образом, во-вторых — наверняка). Как правило, для этого используют мутации, вызывающие врожденные патологии — «менделевские» (наследующиеся в соответствии с законами Менделя) наследственные болезни. Они наследуются по Менделю, потому что вызываются единичными мутациями, а не хитрыми комбинациями десятков и сотен «аллелей риска» в сочетании с факторами среды. В одном из недавних исследований, основанных на этом подходе, темп однонуклеотидных замен у людей был оценен в 1,28 мутаций на 100 млн пар оснований за поколение (1,28 × 10–8 на нуклеотид за поколение) (M. Lynch, 2010. Rate, molecular spectrum, and consequences of human mutation). Поскольку в диплоидном геноме человека примерно 6 млрд пар оснований, это соответствует 77 новым мутациям на геном за поколение.
Другой подход основан на сравнении геномов человека и других приматов. Подсчитав различия в нейтральных (не находящихся под действием отбора) участках генома, можно сопоставить полученный результат со временем жизни последнего общего предка сравниваемых видов (насколько его можно оценить по палеонтологическим данным). Согласно нейтральной теории молекулярной эволюции, скорость накопления нейтральных генетических различий между видами в идеале должна быть просто-напросто равна скорости нейтрального мутагенеза (за одно поколение между двумя видами накапливается столько же различий, сколько новых мутаций возникает у каждой особи). Поэтому, зная время дивергенции, темп мутагенеза можно рассчитать по формуле m = D/2t, где D — число нейтральных различий между видами, t — время жизни последнего общего предка в «поколениях назад». Двойка в формуле появляется из-за того, что оба вида после расхождения накапливали мутации независимо друг от друга.
Впрочем, существует куча факторов, нарушающих правильный ход «молекулярных часов», да и палеонтологические датировки последних общих предков, мягко говоря, не всегда точны. Поэтому надежность данного метода невысока. Неудивительно, что результаты он дал несколько иные. Например, после прочтения генома шимпанзе темп нашего мутагенеза был оценен в 2,2 × 10–8 замен на нуклеотид за поколение, или 132 новых мутации у каждого новорожденного — почти вдвое больше, чем показал анализ наследственных болезней (The Chimpanzee Sequencing and Analysis Consortium, 2005. Initial sequence of the chimpanzee genome and comparison with the human genome).
В последние годы благодаря резкому удешевлению полногеномного анализа появилась возможность оценивать темп мутагенеза напрямую, просто сравнивая геномы родителей с геномами их детей и подсчитывая новые мутации. Появились и другие подходы, в частности основанные на палеогенетических данных. Например, прочтение геномов неандертальцев и других древних людей позволило оценивать темп мутагенеза по числу «недостающих мутаций» в этих геномах, то есть по тому, насколько меньше генетических отличий от общего предка успела накопить линия, вымершая десятки тысяч лет назад, по сравнению с нами сегодняшними. Эти и другие новые методы дают оценки в диапазоне от 1,0 × 10–8 до 1,2 × 10–8 замен на нуклеотид за поколение, то есть 60–72 новых мутаций у каждого новорожденного. И это, по-видимому, близко к истине.
Конечно, это средние значения: у отдельных индивидов число новых мутаций может быть как меньше, так и существенно больше среднего. В любом случае сомневаться не приходится: мы все — закоренелые мутанты! Далеко до нас всякой мелочи вроде бактерий или дрожжей, у которых одна-единственная новая мутация может приходиться на тысячу, а то и на десять тысяч «новорожденных» (клеточных делений) (рис. 1).
У млекопитающих «под отбором» находится 5–10% генома, а всё остальное — в основном мусор (выражаясь политкорректно, это области, все или почти все мутации в которых являются нейтральными, то есть не влияют на приспособленность и не подвергаются действию отбора) (см.: Сравнение геномов 29 млекопитающих проливает свет на эволюцию человека, «Элементы», 05.11.2011). Следовательно, из 60–70 новых мутаций в геноме среднестатистического новорожденного примерно 3–7 являются вредными. Темп возникновения полезных мутаций точно не известен, но они наверняка достаточно редки, чтобы ими можно было пренебречь, говоря об одном среднестатистическом человеке.
От трех до семи новых вредных мутаций у каждого человека в каждом поколении — это угрожающе много. Нужен сильный очищающий отбор, чтобы избежать вырождения, то есть неуклонного накопления генетического груза (см. Genetic load). Если же на отбор надежда плохая, то остается надеяться только на высокие биотехнологии: генную инженерию, генную терапию, экстракорпоральное оплодотворение с искусственным отбором эмбрионов и тому подобное (см.: Алексей Кондрашов, Надежда Маркина. Жизнь без отбора: благо или опасность?). Впрочем, об угрозе вырождения в обсуждаемой статье речь не идет (но эта проблема обсуждается в статьях, на которые ссылаются авторы обзора).
Кроме однонуклеотидных замен бывают еще вставки и выпадения («инделы», см. Indel), инверсии (повороты на 180°) и дупликации фрагментов ДНК разной длины. Такие мутации происходят реже, чем однонуклеотидные замены, зато они затрагивают большее число нуклеотидов и, конечно, тоже влияют на вероятность развития всевозможных болезней. По имеющимся оценкам, пока еще не очень точным, каждый человек несет в среднем около трех новых мелких (1–20 пар оснований) вставок и делеций и 0,16 более крупных (>20 пар оснований).
Зная темп мутагенеза, численность населения и рождаемость, можно примерно оценить общий масштаб генетического полиморфизма современного человечества. Масштаб этот впечатляет: только за время жизни одного последнего поколения в человеческой популяции должно было заново появиться более 1011 точечных мутаций — много больше, чем нуклеотидов в геноме! По-видимому, каждая возможная точечная мутация (кроме несовместимых с жизнью) имеется в данный момент как минимум у сотни-другой живущих на планете людей. Зарегистрированных полиморфизмов, разумеется, гораздо меньше, ведь до поголовного прочтения геномов дело еще не дошло.
2. Закономерности распределения мутаций по геному. Как известно, мутации случайны. По крайней мере, в первом приближении. Это, впрочем, не значит, что вероятность возникновения всех мутаций абсолютно одинакова или что процесс мутагенеза полностью хаотичен во всех своих аспектах. Под «случайностью» мутаций подразумевается вполне конкретная вещь, а именно отсутствие прямого влияния полезности или вредности мутации на вероятность ее возникновения. У живых существ нет механизма, позволяющего рассчитать, какая именно мутация будет им полезна в данных условиях, и внести именно эту мутацию в свой геном. Правда, есть механизмы, позволяющие несколько повысить вероятность появления полезных мутаций (пример — соматическое гипермутирование иммуноглобулиновых генов в лимфоцитах, см.: Мутагенез в лимфоцитах — результат целенаправленного изменения ДНК и последующей «неточной починки», «Элементы», 03.09.2007) и снизить вероятность появления вредных. В частности, оказалось, что частота мутирования у людей связана с временной последовательностью репликации хромосом (см. Replication timing). В участках, реплицирующихся раньше других, возникает меньше мутаций, чем в участках, реплицирующихся в последнюю очередь. Это выгодно, поскольку первыми, как правило, реплицируются участки, в которых много генов. Соответственно, мутации в этих регионах часто оказываются вредными. Последними же реплицируются участки ДНК, в которых преобладает «мусор» и в которых поэтому большинство мутаций оказываются нейтральными.
Наиболее сильный «перекос» в распределении мутаций в человеческом геноме состоит в том, что чаще всего мутируют нуклеотиды Ц (цитозины), за которыми следует нуклеотид Г (гуанин) (см. CpG site). Цитозин вообще является «слабым звеном» в ДНК, поскольку он склонен превращаться в урацил (У) в результате спонтанного деаминирования. Однако системы репарации бдительно следят за тем, чтобы никаких урацилов в ДНК не было, и быстро исправляют большинство возникших таким способом мутаций.
Динуклеотиды ЦГ отличаются тем, что входящие в их состав цитозины часто подвергаются метилированию (см.Метилирование ДНК). Метилированный цитозин в результате деаминирования превращается уже не в урацил, а в тимин — «законное» основание, в норме входящее в состав ДНК. Обнаружить такую мутацию системам репарации гораздо труднее.
В результате частота мутирования цитозинов в динуклеотидах ЦГ примерно в 10 раз выше нормы. Это, в свою очередь, приводит к неравной встречаемости аминокислотных замен. Из всех 20 аминокислот в человеческих белках самым «уязвимым» является аргинин. Как выяснилось, свыше 16% всех аминокислотных замен, приводящих к наследственным болезням, — это замены аргинина (рис. 2). Это факт, на первый взгляд загадочный, объясняется очень просто. Если посмотреть в таблицу генетического кода, то можно убедиться, что все 4 кодона, начинающиеся со склонного к мутациям динуклеотида ЦГ, кодируют именно аргинин.

Рис. 2. Относительная частота аминокислотных замен у человека. Рекордсменом по числу замен является аргинин (R), потому что 4 из 6 кодирующих его триплетов начинаются с динуклеотида ЦГ (данные основаны на анализе 4000 несинонимичных мутаций, вызывающих наследственные болезни). Рисунок из обсуждаемой статьи вScience.
|
В кодирующих последовательностях динуклеотидов ЦГ больше, чем в среднем по геному, что способствует более высокому темпу мутирования кодирующих последовательностей по сравнению с некодирующими. Впрочем, есть также механизм, снижающий частоту мутирования кодирующих последовательностей, по крайней мере тех, которые часто транскрибируются. Дело в том, что транскрибируемые последовательности подвергаются более качественной репарации (см. Transcription coupled repair). Из-за этого в генах, активных в клетках зародышевой линии, наследуемые мутации возникают реже, чем в среднем по геному.
Очевидно, что паттерн мутирования, как и другие признаки организма, вполне может эволюционировать под действием мутаций, отбора и дрейфа. Он может немного различаться не только у человека и шимпанзе, но даже у разных человеческих популяций. И это не только теория. Так, сравнительно недавно, от 40 до 80 тысяч лет назад, изменился паттерн мутирования у предков нынешних европейцев, которые тогда как раз отделились от предков азиатов. А именно, у европейцев повысился темп возникновения мутаций в тринуклеотидах ТЦЦ. Эти тринуклеотиды стали чаще превращаться в ТТЦ (5'-ТЦЦ-3' → 5'-ТТЦ-3') (K. Harris, 2015. Evidence for recent, population-specific evolution of the human mutation rate).
Известно, что именно такие мутации чаще всего возникают в клетках кожи под действием ультрафиолета. Особенно характерны они для клеток меланомы. У европейцев кожа в ходе эволюции стала прозрачнее для ультрафиолета, чем у других человеческих популяций, поэтому учащение подобных мутаций в клетках кожи легко объяснимо. Но вот как эти мутации проникают в зародышевую линию, точно не известно. Одна из гипотетических возможностей состоит в том, что ультрафиолет повышает частоту мутаций данного типа и в коже, и в половых клетках одним и тем же косвенным путем, способствуя деградации фолиевой кислоты. Дефицит этого витамина может приводить к сбоям в ходе синтеза ДНК. Так или иначе, данный факт наглядно показывает, что паттерн мутирования в человеческих популяциях действительно подвержен эволюционным изменениям.
3. Немолодые отцы — главный источник наследственных мутаций. На сегодняшний день твердо установлено, что львиную долю новых наследственных мутаций люди получают от отцов. При этом чем старше мужчина, тем больше мутаций в его сперматозоидах (о причинах этого рассказано в новости У шимпанзе, как и у людей, число мутаций у потомства зависит от возраста отца, «Элементы», 18.06.2014). Около 95% вариабельности потомков по числу новых мутаций объясняется возрастом отца. Хуже того, оказалось, что у старых отцов сглаживается упомянутая выше зависимость частоты мутаций от последовательности репликации (replication timing). Соответственно, растет доля мутаций в «осмысленных» участках генома, а среди таких мутаций выше доля вредных.
С возрастом матери число мутаций в ее яйцеклетках не увеличивается, зато вероятность рождения детей с хромосомными нарушениями, такими как синдром Дауна, растет.
Короче говоря, заводить детей все-таки лучше в молодости. Наблюдающаяся в развитых странах тенденция к увеличению среднего возраста отцовства и материнства дополнительно усиливает риск генетического вырождения человечества.
4. Соматические мутации и их медицинское значение. За жизнь человека клетки его тела делятся триллионы раз. Каждое деление сопряжено с риском соматических мутаций, да и в промежутках между репликациями молекулы ДНК могут повреждаться. В тканях, клетки которых делятся особенно интенсивно (например, в кишечном эпителии), к 60 годам должна присутствовать, хотя бы в одной клетке, едва ли не каждая из всех возможных точечных мутаций. Разнообразие соматических мутаций выше, чем наследственных, потому что первым, чтобы не быть немедленно отбракованными отбором, достаточно быть совместимыми с жизнью всего-навсего одной клетки, тогда как вторым необходима совместимость с жизнью целого организма.
Хотя соматические мутации не передаются по наследству, их медицинское значение очень велико. Давно известно, что они играют ключевую роль в развитии различных видов рака (см.: На пути к детальному каталогу раковых генов, «Элементы», 06.04.2015). В последние годы стало ясно, что соматические мутации вызывают и многие другие заболевания (R. P. Erickson, 2010. Somatic gene mutation and human disease other than cancer: An update). Например, выяснилось, что соматические мутации в генах PIK3CA, AKT3 и mTOR вызываютгемимегалэнцефалию — одностороннее увеличение и нарушение функции одного из полушарий мозга, которое к тому же повышает риск развития эпилепсии. Сравнительно небольшая доля мутантных клеток может нарушить работу обширных областей коры: у пациентов с дисфункцией целого полушария мутацию могут нести лишь от 8 до 35% клеток мозга. По-видимому, соматические мутации лежат в основе многих других патологий центральной нервной системы (A. Poduri et al., 2013. Somatic Mutation, Genomic Variation, and Neurological Disease).
5. На пути к пониманию фенотипических эффектов мутаций. Конечной — и едва ли достижимой в обозримом будущем — целью изучения человеческих мутаций является создание их полного каталога с указанием влияния каждой мутации на фенотип. В идеале хорошо бы еще разобраться во взаимном влиянии эффектов разных мутаций друг на друга (см. Epistasis), но до полного каталога таких взаимодействий пока еще слишком далеко.
Эффекты мутаций изучают на трех уровнях, которые можно условно назвать молекулярным, медицинским и эволюционным. В первом случае речь идет о том, как та или иная мутация влияет на экспрессию гена или функцию белка. Во втором — о том, как мутация влияет на вероятность развития тех или иных заболеваний. В третьем — о влиянии мутаций на приспособленность (репродуктивный успех). Конечно, это взаимосвязанные вещи, но корреляции между ними не строгие. Например, мутации, вызывающие старческие болезни, вряд ли будут «вредными» с эволюционной точки зрения: они не скажутся на репродуктивном успехе. Ослабление функции какого-нибудь фермента может сказаться на здоровье человека в одних условиях среды, но никак не проявиться в других, и так далее. Все три типа исследований связаны с большими методологическими трудностями, поэтому пока мы имеем более или менее детальную информацию лишь по относительно небольшому числу мутаций.
Например, выяснение молекулярных эффектов мутаций — это чрезвычайно кропотливая работа, которую, однако, можно проводить в пробирке. В итоге, потратив неимоверное количество сил и средств, можно получить более или менее полный мутационно-функциональный спектр для какого-нибудь белка. На рис. 3 показаны результаты исследования влияния разных аминокислотных замен на убиквитин-лигазную функцию (см. Ubiquitin ligase) регуляторного белка BRCA1. Это белок, пришивая убиквитин к другим белкам, регулирует репарацию ДНК и играет важную роль в защите от рака.

Рис. 3. Рисунок показывает, как влияют на убиквитин-лигазную функцию BRCA1 аминокислотные замены в каждой из 103 аминокислотных позиций исследованного фрагмента белка. Аминокислотные позиции расположены вдоль горизонтальной оси и подписаны в нижней части диаграммы. Для каждой позиции разными цветамипоказаны эффекты разных замен. По вертикальной оси отложены 20 аминокислот, которыми исследователи поочередно заменяли исходную аминокислоту в каждой позиции. Желтыми прямоугольничками отмечены аминокислоты «дикого типа», то есть те, которые стоят в данной позиции в нормальном, не мутантном белке. Синий цветобозначает ослабление функции, красный — ее усиление выше нормального уровня,белый — сохранение исходного уровня активности белка (см. цветовую шкалу справа; единица соответствует исходному состоянию, то есть «норме»). Наконец, серый цветобозначает, что для данной замены данные не были получены. По количеству белых ипочти белых прямоугольничков можно судить о толерантности данного белка к мутациям, то есть о спектре несинонимичных (ведущих к замене аминокислоты) мутаций, которые не приводят к драматическим изменениям его функциональности. Изображение из статьи L. M. Starita et al., 2015. Massively parallel functional analysis of BRCA1 RING domain variants
|
Данное исследование, с одной стороны, впечатляет своей грандиозностью, с другой — наводит на грустные мысли о том, сколько же сил нужно потратить, чтобы получить хотя бы такие простенькие, одношаговые «адаптивные ландшафты» (см. Fitness landscape) для всех человеческих белков и всех единичных несинонимичных мутаций, не говоря уж об их комбинациях. Но тем не менее эта работа необходима, если мы хотим приблизиться к ответу на ключевой вопрос о соотношении генотипа и фенотипа. Без такого знания мы не сможем разработать технологии, которые позволят в будущем заменить слабеющий естественный отбор целенаправленным редактированием генома, остановить накопление генетического груза и даже, может быть, улучшить человеческую природу.
Подписаться на:
Комментарии (Atom)